
Sakharov D.,
The R_E.Kavetsky Institute of Experimental Pathology, Oncology and Radiobiology,
National Academy of Science of Ukraine
Waiserman A., Koshel N.,
The Institute of Gerontology, National Academy of Science of Ukraine
Gabor Lednyiczky,
Ffippocampus Research Facilities, Central Division,
1031 Budapest, Ninesi tat 67, Hungary, let/fax 36-1-1886865
ABSTRACT:
It has been long suspected that biological electromagnetic waves may play an important regulatory role in life of living systems. To demonstrate this phenomenon, we tried to exert control over the viability and stress-resistance of flies (which have undergone an activating and depressing influence of a temperature) by transferring the biological electromagnetic waves from one group of bio-objects to another. The considerable improvement of the viability indices’ and stress-resistance of the heat-shock depressed Drosophila melonogaster flies until normal values returned. Moreover, the influence of the waves under certain conditions is able to optimize the system’s function activity of the heat-shock activated and heat-intact flies.
INTRODUCTION
In reviewing the voluminous literature on the electromagnetic phenomena of living systems, one can find a strong correlation between the dynamics of different physiological processes and characteristics of endogenous electromagnetic signals or fields (EMFs). Taking into account the connection between a wide range of electromagnetic waves and the state of living systems, one can draw two explicit conclusions about the role that such waves may play: (I) pointing to a certain functional status of a system; (2) promoting or inhibiting specific processes.
To verify the second conclusion practically, one needs to induce remarkable changes in the blo-object under investigation by means of biological EMFs. Though many scientists corroborated the possible regulatory role of intrinsic EMEs by indirectly [Weyer R A., 1985; Choy ft et al., 1987] exposing an organism to artificial EMFs, Gunvitch was the first one who did it using its natural biological EMF’s [Gurvvitsch A.G., 1932]. One further example of similar experiments is the work recently done by Dr. Ho M.W. [Ho MM., N1992]. She reports that even a brief exposure of Drosophila’s embryos to external weak magnetic fields leads to crucial abnormalities in the pattern of body formation in the larvae which hatch twenty four hours later. It is difficult to explain the mechanisms that are responsible for that effect. However, one would not neglect the possibility of interference between biological and applied EMFs resulting in disorder of the proper influence of intrinsic EMF’s on the major pattern determination processes.
These considerations in mind, we supposed that intrinsic EMFs actually play a regulatory role in the determination process and are important in intercellular communication. In order to practically confirm some of these assumptions, we used the “BICOM” – device as a suitable tool for the electromagnetic information transfer between objects and chrysalises of Drosophila melazzogaster as an available object of investigation with a high degree of the proliferation and determination cell activity [Lints E.,1985; Mayer P.,1985]. Drosciphila melanogaster, or fruit fly, has been extensively used in genetic investigations and is probably the best understood animal in terms of inheritance studies_
It is well known that a 40°C heat shock may considerably decrease the fly’s viability, i.e. its capacity to tolerate future stress conditions: a 30% mortality in the embryo population, and evocation of some abnormalities during development of survived embryos. On the molecular level, it is reflected as a sharp changes in the spectrum of gene activity the functional activity of 90% of genes are suppressed but not those responsible for heat shock proteins production [Craig E.,1985].
We have recently shown the possibility of significantly invigorating the viability of chrysalises and deriving from them flies by heat treatment with certain parameters (120 min:, 37°C). This heat shock activation of vitality may be considered a reaction of non-specific adaptation of the organism [Garkavi L. et al, 1990]. With respect to this, the main task of the experiment was to correct the temperature-induced considerable deviations from the normal development in the Drosophila’s chrysalises by means of the endogenous EMFs originating from the heat shock activated chrysalises.
MATERIALS AND METHODS
The outbreed Oregon- R wild type laboratory population of Drosophila melanogaster was used in the experiments. The embryos were placed in one-liter glass vessels containing a standard medium composed of sugar, agar, dried yeast, semolina porridge, and mold inhibitor’ . They were constantly kept at 25° C, the humidity of the experimental cabinets was 40-60% RI-I, Proper administration of light (12 hours of light and 12 hours of darkness) was employed.
On the stage, chrysalis flies underwent the influence of the two different heat shock regimes: a) 120 min. at 37°C – the “activating” heat shock regime; b) 120 min. at 40°C – the “depressing” heat shock regime..In connection with this we have included into the experiment next groups: “N” groups consisted of the temperature unaltered chrysalises; “A” groups consisted of the temperature-activated ones and “D” denoted the groups of temperature-depressed chrysalises.
Then the groups were divided into experimental subgroups depending on the quality of electromagnetic information they experienced. On average, every subgroup consisted of >1000 chrysalises that continued to live in one-liter glass vessels.
The BICOM device (Brilgelnann GmbH, Germany) was used to acquire the electromagnetic field of the chrysalises situated on the input applicator, to modify these fields and transduce them onto the experimental group of chrysalises placed at the output applicator,
Hereafter, the programs of treatment is denoted as follows: The logic program occupies the Position 1 (amplification or attenuation), Position 2 scans frequencies through the whole band-pass of the filter provided that narrow frequency widows are opened in tandem, Position 3 controls the speed of scanning, Position 4 controls the interval or continuos operation, Position 5, the duration of treatment, and Position 6.
- H, 0.9, Frequency run -15″, Continuos 20 sec.
- 0.9, Frequency run -15″, Continuos 60 sec.
- H, 0.9, Frequency run -15″, Continuos 30 min.
Based on the results of the former experiment dealing with the effects of endogenous Elvin on the imago of Drosophila (Tab. 1), the two first programs were shown to be the most effective ones. The :last one is chosen in order to analyze the dose-dependence or the occurrence of “dose window” [Lawrence A.,1982; Choy R.,1987; Webb S.,19841 of the influence.
The 1-12th (the duration of influence 20 and 60 sec) experimental groups experienced the influence of endogenous ELF-ELI EMFs two days four times daily with a one hour interruption between treatment sessions. The 13-15th chrysalises groups duration of influence was 30 min. for two days, once daily.
To generalize the results of the both applied influences – heat shock and EMFs – in terms of the vitality of the chrysalises and flies, we analyzed several representative indices.
The percentage of mortality of chrysalises, i.e. the volume of chrysalises that did not finish metamorphosis successfully.
To present the qualitative analysis of the fecundity of the young females, i.e. the mean number of eggs produced by 100 females from every subgroup during a day, they were separated from males into 200 ml vessels on the third day after the emergence2 . Fecundity of flies is considered normal if the eggs undergo complete metamorphosis and is denoted in the tables below by “plus”, the successful embryonic development of approximately a half of all produced eggs can be considered reduced fecundity and is denoted by-“plusiminus”. When produced eggs turn to be non-viable, that is almost no.. fees: appeared from .them one can say this is completely depressed fecundity – it is denoted by “minus”.
In order to analyze the index of mean life-span (MIS) only males from every experimental group – 125 randomly chosen flies – are placed into five special vials of 25 flies each. The diameter of the vial is 10 mm and length – 150 mm. All these vials were also supplied with 2 ml of standard medium. Transfers to a fresh medium were made 3 times a week until the complete elimination of the experimental populations. The mortality of the flies is estimated by daily performed observations of vials with counting the dead Drosophila ts. On the basis of this data, the mean life span was calculated for every experimental group.
The time of movement (sec) of the flies towards a source of directional light stimulus, the positive phototaxis, was controlled since this index directly correlates with integral viability of an organism [151 The flies of a certain experimental group were placed into a 10 mm x 500 .rnrn glass tube with the source of light, a 100 W electric bulb, situated on the distant edge of the tube. We measured the time in seconds for 50% of the population to transfer from the zero point to a mark 100mm away.
The resistance of flies to heat shock (min.) is evaluated in terms of the time it takes for the death of 50 % of population by heating to 40°C. For this propose, males are placed in 5 vial q free of medium, 20 flies to a vial:
To estimate the resistance of flies to starvation (hour), 100 flies are maintained in 5 vials without medium and the time taken to complete a 50% mortality of the population at 22°C is assessed.
Mathematical analysis of the data was performed by using “Statgraphics” software [ 16]
RESULTS AND DISCUSSION
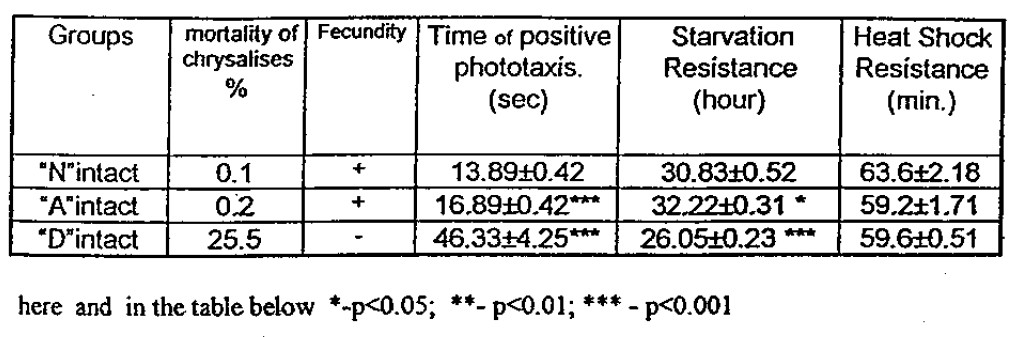
As it can be seen from Table 2., the ‘A’ and D’ administration of heat shock influences the viability of chrysalises and young flies variously. Only the values of the Positive Phototaxis (PP) – (the time of movement to the label) were of the same type. The locomotor activity of flies was decreased in both cases. In the case of D’ administration of HS the depression was profoundly expressed. At the same time, the resistance of flies to starvation was the opposite. The flies of “A” intact group (intact with respect to the information transfer by BICOM ) appeared to be much stronger in their resistance to starvation. Thus We can draw the conclusion that the administrations affected the flies in a • different way: If the “D” regime only decreases integral viability of flies then “A” redistributes the potential resources of viability to provide a proper “answer” of organism while it shifts from normal to unusual (starvation, any stress response) living conditions. Likewise, such redistribution occurs on the level of genes. It is well known that while heat shock takes place 90% of synthesized proteins are the HS-proteins and the synthesis of the other is suppressed [6]
Besides the “D” temperature regime considerably disturbs the metamorphosis and cause the high mortality (25.5%) of chrysalises (the stage after larvae) and completely suppressed the fecundity of young females.
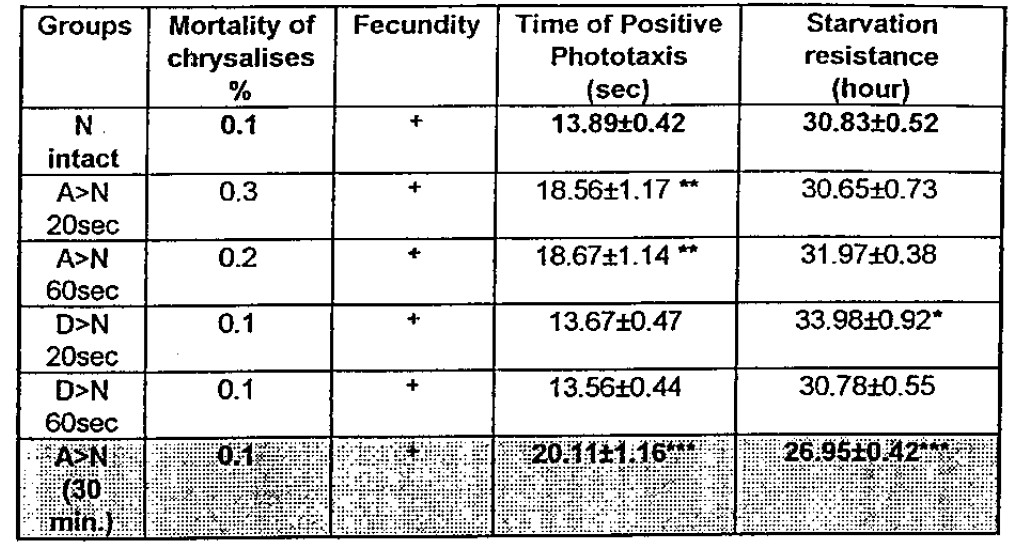
In three of five trials, the influence-of the Elias induced a significant decrease in the speed of movement of flies from heat-shock intact (so called ‘normal”) subgroups. In the group A>N (duration of -treatment 30 min.) the starvation resistance of flies is decreased as w911 as their mobility. In other words, this program brings about a total decrease of viability (overdosing). It is difficult to explain why the result of the D>n (20 sec) regime showed an increased in the SR without any changes in mobility.
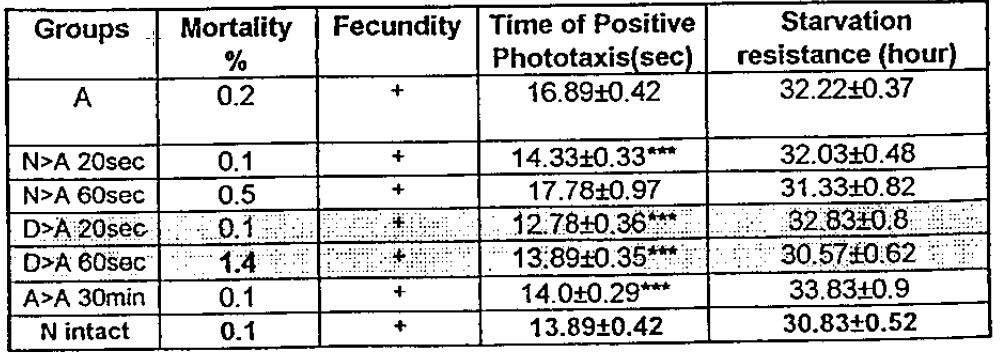
Considering the information transfer from the population of the flies of subgroups “N” or “D” to the “A” flies (Table 4,), one may notice an interesting peculiarity in the results. In four of five trials, the influence resulted in the significant normalization of mobility of flies returning to the value of intact normal flies (see. Table 1.) at the same time retaining the positive effect of the 37°C heat shock on the starvation resistance of the flies. Thus it is possible to prepare an organism to fight against stress conditions while avoiding the negative effects of non-specific adaptation (or the above mentioned redistribution of the life resources).
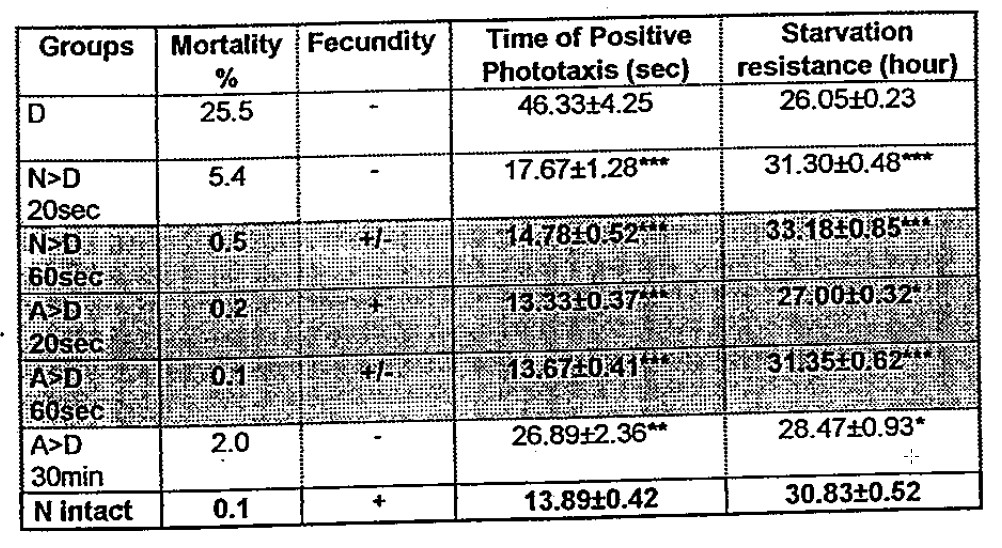
The effects of the e.m. influence turned to be most pronounced when the temperature-depressed subgroups were treated as shown in Table 5. In all trials, the mobility and resistance to starvation was so sharply increased that it shows no difference from that of normal intact (pure control) flies and chrysalises. Furthermore, we were able to completely cease the mortality of chrysalises in the trials N>D 60 sec., A>D 20 sec., A>D 60 sec., A>D 30 min., and slightly in N>D 20 sec. Nearly the same results appear in the restoration of the females fecundity (completely restored in A>D 20 sec.). It worth mentioning that significant positive shifts in the viability’s indices of flies was observed provided that the source of e.m. signals was the group of heat-shock depressed flies (see D>A and D>N modes in Table 3, 4). A clarification of this point would require more profound theoretical and practical approaches.
CONCLUSION
The electromagnetic waves that are irradiated by an organism under certain conditions can cause significant modifications to the functional activity of another organism’s system. As such, these waves not only play a role in the regulation of an organism’s functions, but they also can be involved in the regulation of a whole population of organisms such as the populations of Drosophila flies or Daphnia that was discussed in [Calle M,1992].
Since weak, biologic.ally meaningful e.m. signals were successfully accepted by the population of-flies-in their environments wrenagainst much greater background EMFs (usually to be found inside buildings), the results suggest the existence of the “natural selectivity” of living systems.
Thus we conclude a possible correction of the heat-induced damages and in all probability, any pathological changes by means of information transfer from bioobjects having the earlier acquired mechanism of stress-resistance.
The following part of the discussion covers the explanation of possible genetic mechanisms that can be responsible for the observed results. According to the NeoDarwinsitic explanation of evolution, the major changes in evolution arise from the accumulation of point mutations in regulatory and structural genes which then undergo, the pressure of natural selection and can therefore be eliminated or stabilized. The stabilization of acquired characters occurred first of all as the inheritance of the structural peculiarities of genes instead of a pattern of their functional activity. The latter is possible only if the existence of La.marckian inheritance is considered [Surani M.,I994]. It is relevant to question also how genetic events that can be observed in somatic cells relate to the variations in organization of germline DNA. According to the concept of the Weismann’s barrier [Weisman, 1893], the soma and germline are distinct and separate. Thus, the germplasm is immune from both the influence of events taking place in sorna and environmental influences such as stress, nutritional status, etc. However, as it is being recognized today, genomic organization and the relationship between somatic and germline genes is infinitely more complex and much more susceptible to dramatic changes than previously considered. In light of the facts excellently reported and _ analyzed: in, [Pollard J. W., 1988] the dogma of germline sequestration (isolation) seems to be wrong. The author argues that “if an isolated population of organisms were exposed to an altered environment, rapid variation of genomic organization might occur with concomitant effects on morphology, size and pattern of gene expression”. We suggest the result of the experiment presented in this paper in great extent evidence to the accuracy of the author’s statement since while chrysalises were treated by heat and ElvIFs, the effects were observed on the stage of imago when all cells that underwent the influences were substituted by young ones inherited the acquired pattern of the gene’s activity.
Epigenetic inheritance is possible due to the process of genomic imprinting. A genom fixes in “memory” some epigenetic events and is able, under certain conditions, to pass the imprint to the next generation of cells [Barlow D.,1994; Jablonka E. et al., 19921. On the topic of ontogeny3, one usually considers both processes. In the case when germline cells are involved in the process, a newly-born organism can inherit the acquired imprint from. During the last few years, the data obtained points to the possibility of environmentally evoked alterations in the epigenom [Holliday 8,1990; Sancar A. et 19901_ Simply, it means that besides the inflow of information from DNA to proteins, the opposite direction of the process is also feasible and may give rise to the new inherited status of genom. It was verified in the investigation of the influence of external agents on the phenotype of an adult organism provided that the influence was applied at early stages of embriogenesis (lose 1 et al., 19911 Effects of the influence are accounted for as follows: (1) it alters activity of the embryo’s physiological systems; (2) it causes the accumulation of somatic point mutations; (3) the influence alters the functional activity of genes then the new functional activity is imprinted. However, if the first two suppositions can easily explain the negative effects on phenotype, the third one is the only explanation for positive effects such as the acquired resistance of an organism to stress. Throughout numerous experiments (unpublished data) while larvae or chrysalises4 were exposed to certain doses of heat shock or X-rays, we observed both an increase and decrease in viability of adult Drosophila in. Of course, to prove epigenetic mechanisms of the observed, we would determine the minimum extent of heat-shock protein production in the cells of treated flies during the next experiment.
REFERENCES
- Adhys Sancar, Garges Susan. Positive control II 1. Biol. Chem.-1990.- -265,19.-p.10797-10800
- Barlow D.P. Imprinting: A gamete’s point of view (Trends in Genetics.- 1994.- v. 10.- N6.- p. 194-198)
- Choy R I.S., Morro J.A., Smith C.W. Electrical Sensitivity in Allergy Patients // Clinical Ecology.–1987.–4,3.—p.93-102
- Craig E.A. The heat shock response // Crit. Rew. Biochern.-I985-18.-p239-280
- Galle M. Population density -dependence-of Biophoton Emission from Daphnia II Recent Advances in Biophoton Research and its Applications. Edited by Popp F.A_, Li K.L., and Gu Q.- “World Scientific”- Singapore-New Jersey-London-Hong Kong.- 1992.-p.345-353
- Garkavi Kwakina E.B. Ukolova M.A. Reactions of adaptation and resistance of an organism. – Rostov University Publish House, Rostov-na-Donu, 1990.-223 P. (n Russian)
- Gurwitsch A.G. Die mitogenetische Strahlung der optischen Bahn bei adaquater Erregung i/Pflugers Arch.ges. Physiol.-1932.-231.- p.254-264
- Ho M.W. Xu X, Ross S., Saunders P.T. Light Emission and. Rescattering in Synchronously Developing Population of Early Drosophila Embryos II Recent Advances in Biophoton Research and its Applications. Edited by Popp F.A., Li K.L., and Gu Q.- “World Scientific”- Singapore-New Jersey-London-Hong Kong.-1992.p.287-304
- Ho M•W., Stone TA, Jerman I., Bolton J., Bolton H., Goodwin B_C., Saunders P.T., and Robertson F. Brief exposure to weak static magnetic fields during early embriogenesis causes cuticular pattern abnormalities in Drosophila larvae II Phys. Med. Biol. – 1992 a.-37,5.-p.1171-1179
- Holliday R. Paradoxes between .genetics-and –development II J Cell Sci,- 1990.- v. 97.- Ni- p. 395-398
- Jabionka E., Lachmann M., Lamb M.J. Evidence, mechanisms and models for the inheritance of acquire characters // J. Theor. Biol..- 1992.- v. 158.- N2.- p. 245-268
- Klose 3., Howlet S.K., Barton S.C., Surani M.A., Gurtmann I., Reik W. Adult phenotype in the mouse can depend on epigenetic events in the early embryo: [Abstr.1 2nd Mammal. Genet. and Dev. Workshop, London.- 1991
- Lawrence A.F. Adey W.R. Nonlinear Wave Mechanisms in Interactions between
Excitable Tissue and Electromagnetic Fields // Neurological Research.–I 982.-4,1-2.-p.1 I5-15= - Lints F.A. Insects Handbook of the biology of aging NY, 1985-p146-149
- Mayer P.J., Baker III G,T. Genetic aspects of Drosophila as a model system of eucariotic aging // Intern. Rev. Cytol.- 1985.-95. -p.61-102
- Pollard J.W. New genetic mechanisms and their implication for the formation of new species II Evolutionary Processes-and Metaphors . Ed. by M. W. Ho & S.W. Fox.Chichester-New York-Brisbane-Toronto-Singapore . JOHN WILEY & SONS. -1988.- p.63-85
- Recent Advances in Biophoton Research and its Applications. Edited by Popp F_A., Li K.L., and Cm Q.- “World Scientific”- Singapore-New Jersey-London-Hong Kong.-1992.-504 P.
- Statgraphics.-User’s guide-1987.- vol.1-6
- .Surani M.A. Genomic imprinting: Control of gene expression by epigenetic inheritance II Current Opinion in Cell Biology.- 1994.- v.6.- N3.- p. 390-395.
- Webb Si. Nonlinear phenomena in bioenergetics and oncology as seen in 25 years of research with millimeter microwaves and Raman spectroscopy II Nonlinear Electrodynamics in Biological. Systems. Ed. by Adey W.I. 8c Lawrence A.F.-Plenum Press, London-New York.-1984.-p.549-567
- Weisman A. The Germ Plasm: A Theory of Heredity, Scott Publishing, London.
- Wever R.A. The electromagnetic environment and the circadian rhythms of human subjects .11 Biological Effects and Dosirnetxy of Static and ELF Electromagnetic Fields. Ed. by Grandolfo M., Michaelson S.M., Rindi A. —Plenum Press, New York & London.-1985.—p.514
